
Трилобиты







Фрагменты от 5 трилобитов. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найдено: Ленинградская область, река Волхов.
Трилобит № 31. ЦЕНА - 1500 руб. за все.
Контактный email: 1943info@gmail.com ОФОРМЛЕНИЕ ЗАКАЗА Все предметы, из старой, личной коллекции.





Свернувшийся трилобит. Сохранились глаза. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 30. ЦЕНА - 800 руб.




Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 29. ЦЕНА - 1200 руб.




Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 28. ЦЕНА - 1200 руб.




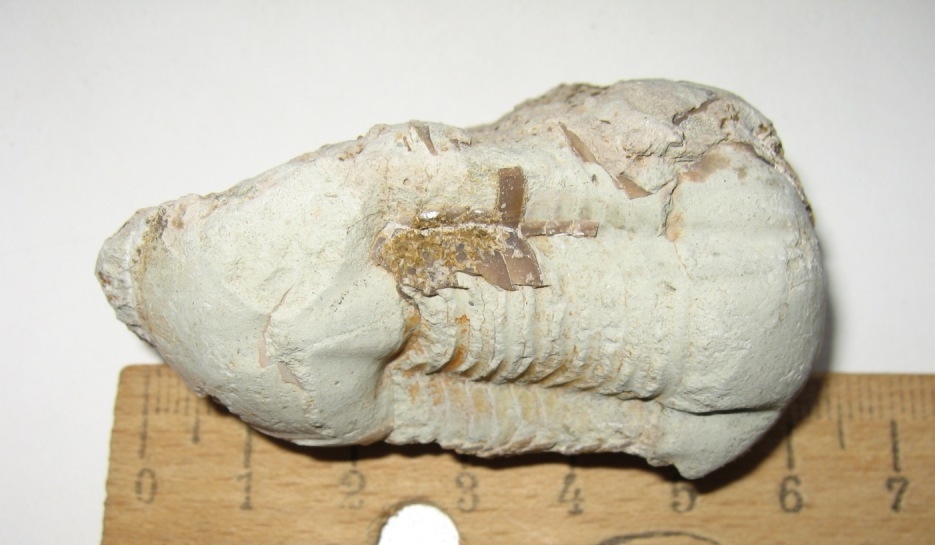
Два крупных фрагмента трилобитов. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найдено: Ленинградская область, река Волхов.
Трилобит № 27. ЦЕНА - 1200 руб. за все.



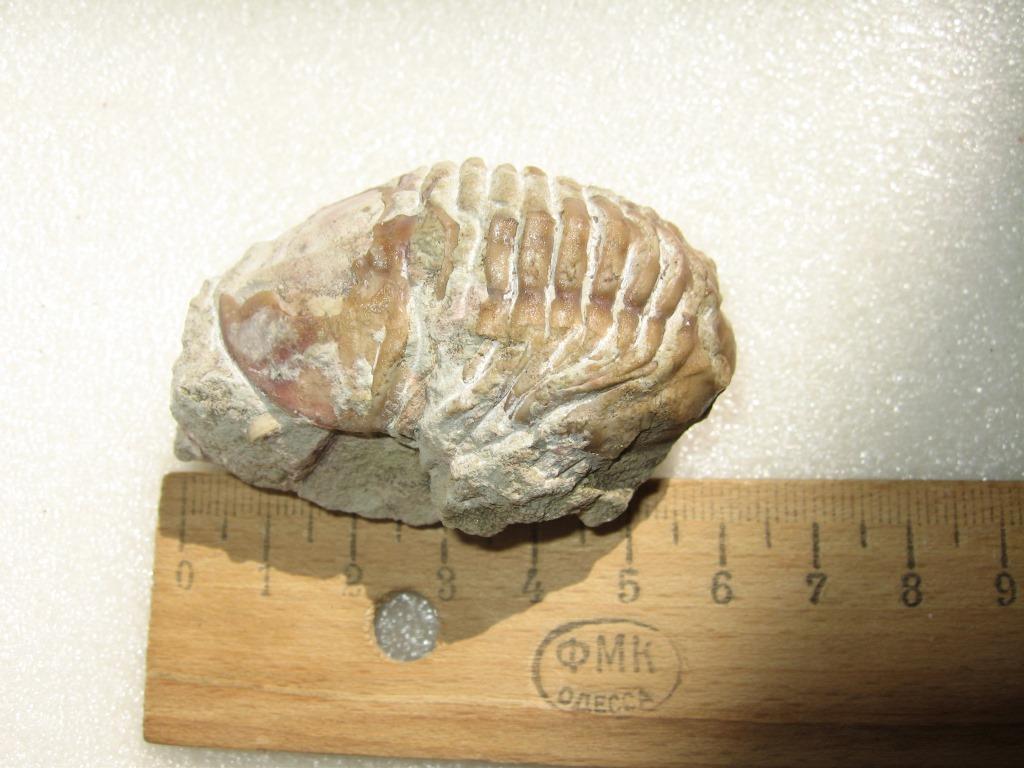
Трилобит. Найден: Ленинградская область, Путилово.
Трилобиты — вымерший класс морских членистоногих. Известно свыше 10 тыс. ископаемых видов и 5 тыс. родов, объединяемых в 150 семейств и 9 отрядов. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад.
Трилобит № 26. ЦЕНА - 800 руб.





Трилобит на породе. В идеальной, коллекционной сохранности! Найден: Марокко. Примерно 450 миллионов лет.
Трилобит № 23. ЦЕНА - 4000 руб.




Свернувшийся трилобит. Крупный. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Подкрашен. Найден: Ленинградская область, река Волхов.
Трилобит № 18. ЦЕНА - 1000 руб.









Трилобит Cybele panderi, с шипами - данный вид, крайне РЕДКО встречается!!! Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов. РАРИТЕТ !
Трилобит № 17. ЦЕНА - 12 000 руб.








Свернувшийся трилобит на породе. Найден: Ленинградская область, Путилово.
Трилобиты — вымерший класс морских членистоногих. Известно свыше 10 тыс. ископаемых видов и 5 тыс. родов, объединяемых в 150 семейств и 9 отрядов. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Отличный экземпляр в коллекцию или в подарок.
Трилобит № 16. ЦЕНА - 4500 руб.








Свернувшийся трилобит. Крупный. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 15. ЦЕНА - 4000 руб.




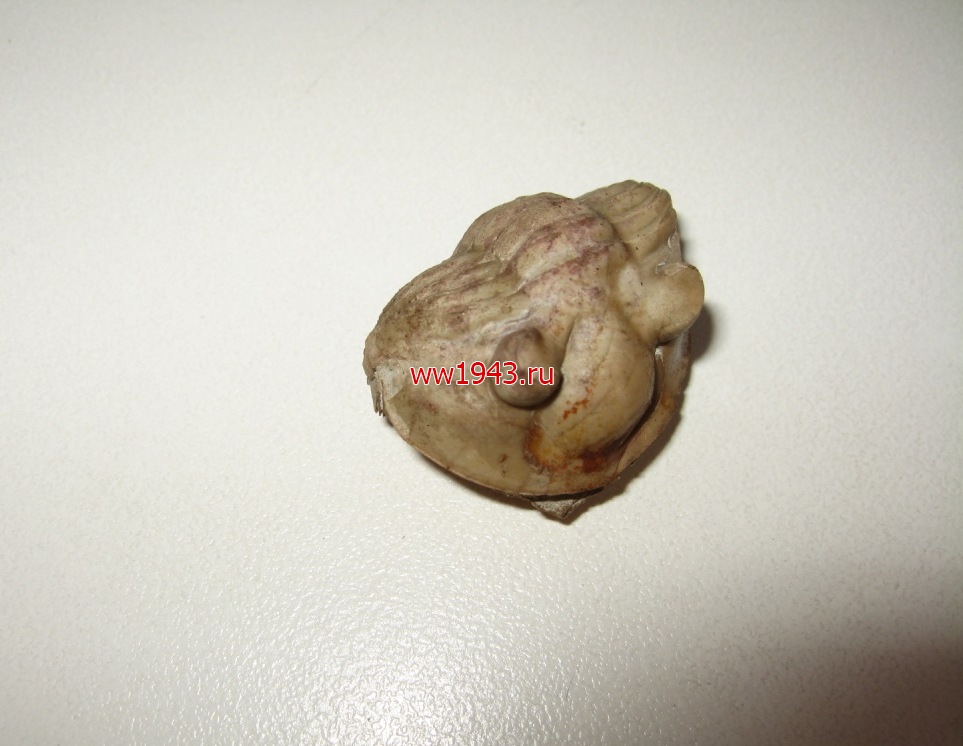
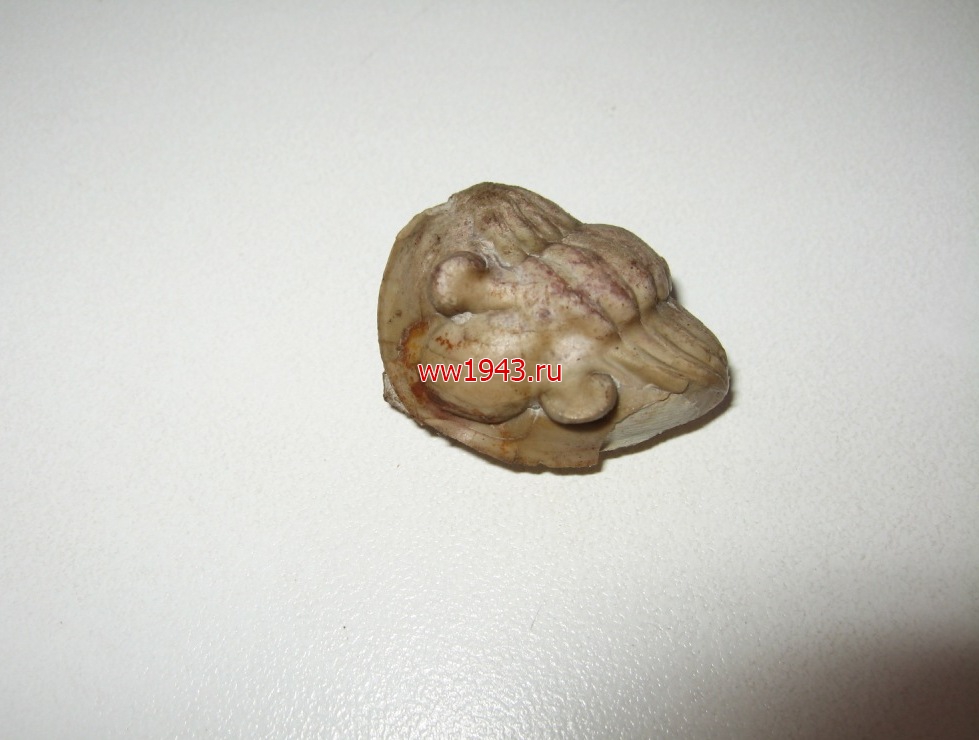
Свернувшийся трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 14. ЦЕНА - 4000 руб.




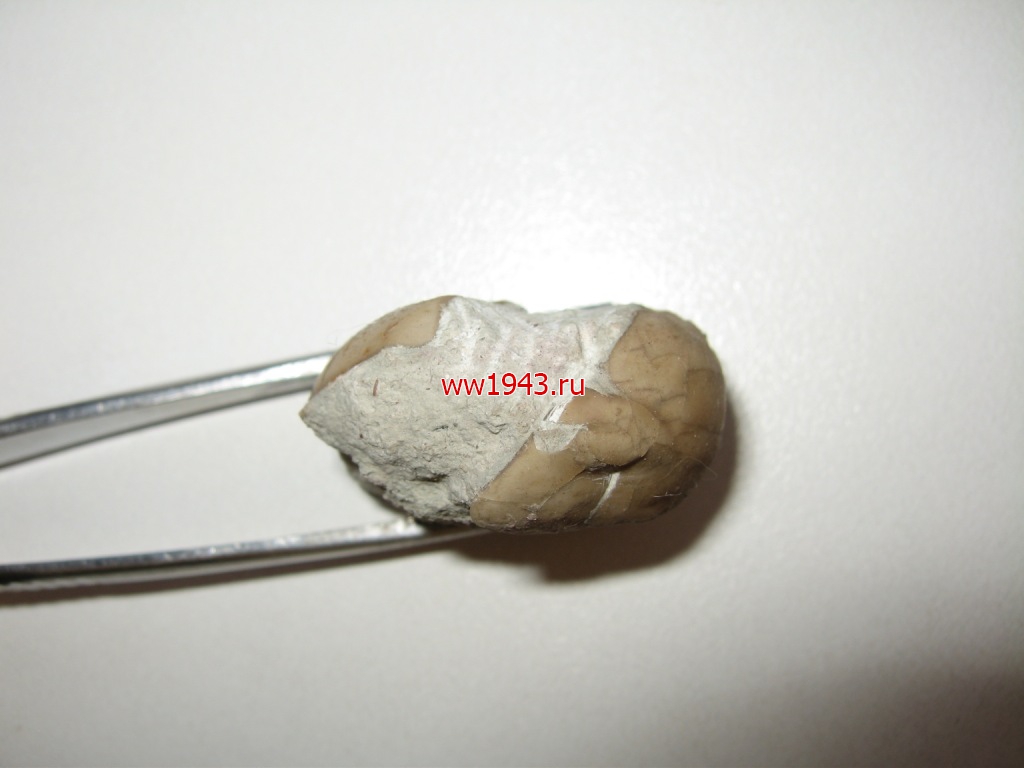

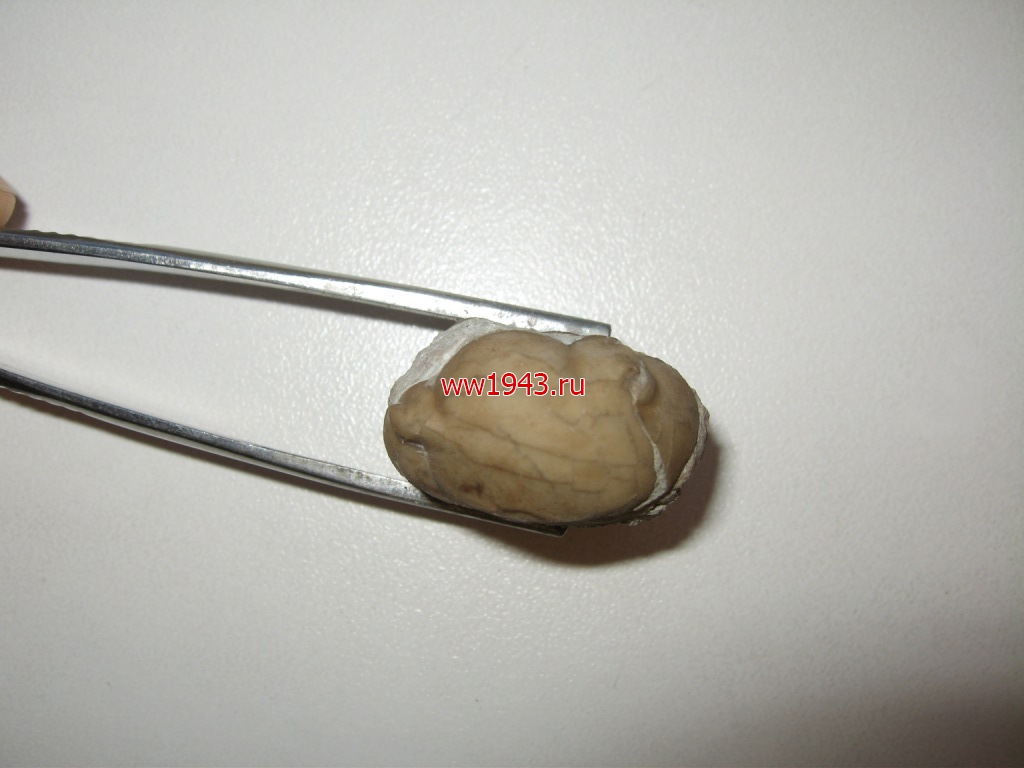

Свернувшийся трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 13. ЦЕНА - 700 руб.




Свернувшийся трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 12. ЦЕНА - 1000 руб.





Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 11. ЦЕНА - 800 руб.





Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 10. ЦЕНА - 1500 руб.







Свернувшийся трилобит. Сохранились глаза. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 9. ЦЕНА - 800 руб.

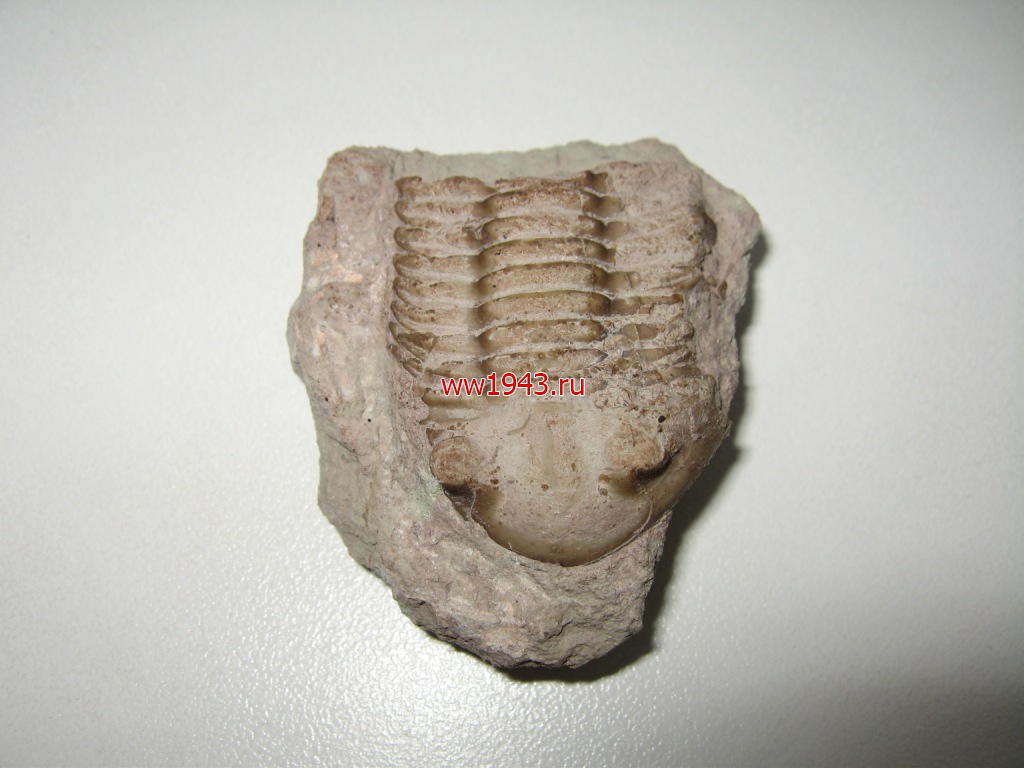


Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 8. ЦЕНА - 1000 руб.





Трилобит. Голова не сохранилась, в остальном, сохранность панциря очень хорошая. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 7. ЦЕНА - 800 руб.





Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 6. ЦЕНА - 700 руб.



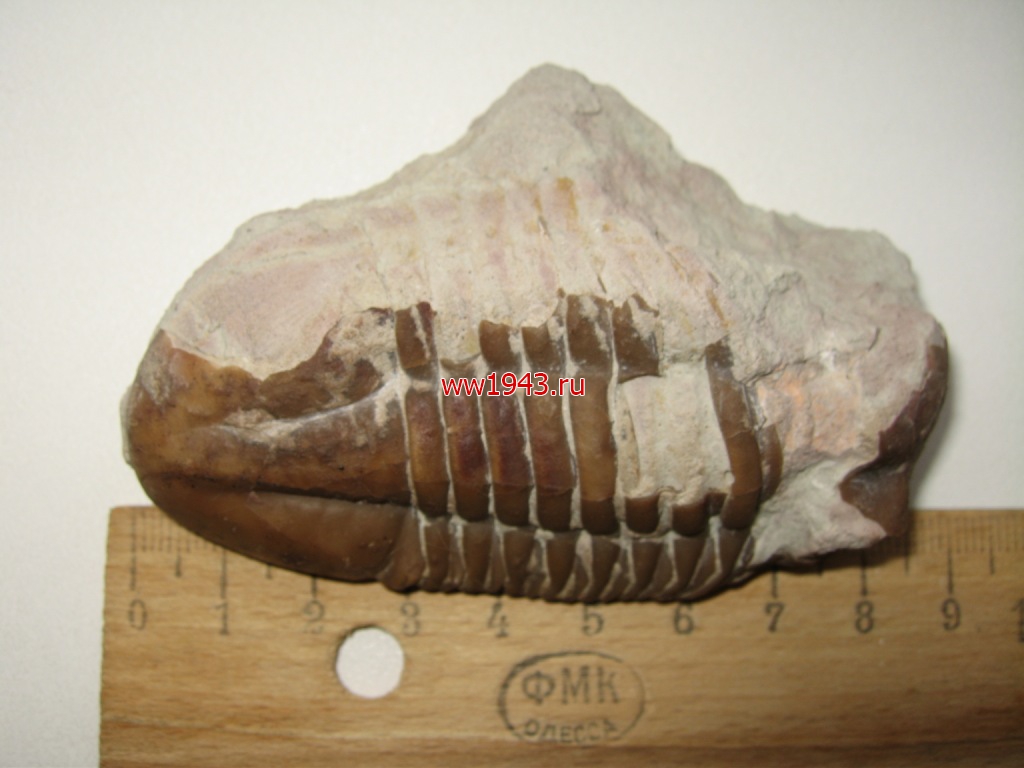

Крупный трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, река Волхов.
Трилобит № 5. ЦЕНА - 500 руб.

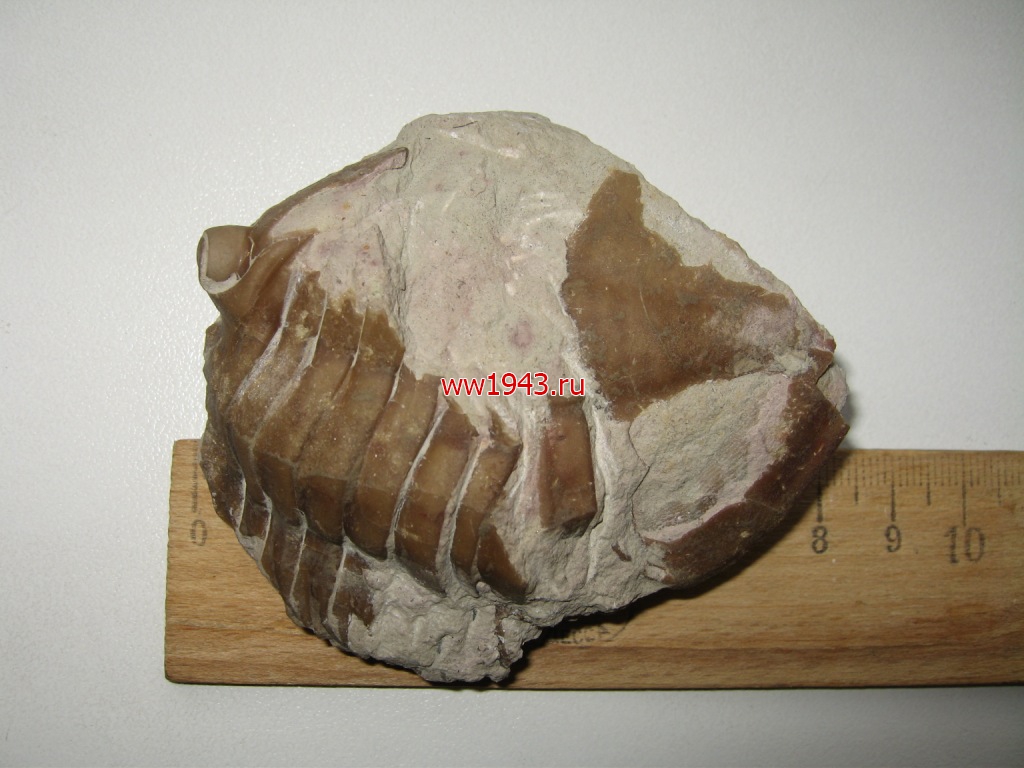




Крупный свернувшийся трилобит. Залегает в известняке немного свернувшись. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден в Ленинградской области, река Волхов.
Трилобит № 4. ЦЕНА - 600 руб.




Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден в Ленинградской области, река Волхов.
Трилобит № 3. ЦЕНА - 300 руб.




Свернувшийся трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден в Ленинградской области, река Волхов.
Трилобит № 2. ЦЕНА - 200 руб.




Трилобит. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден в Ленинградской области, река Волхов.
Трилобит № 1. ЦЕНА - 400 руб.

Трилобиты
Временный диапазон: 521-252 млн. Лет
Ранний кембрийский - поздний пермский
Kainops invius боковые и вентральные
Kainops invius, ранний девон
Paradoxides
Paradoxides sp., Поздний кембрийский
Научная классификация
Королевство: Animalia
Тип: Членистоногие
Подтип: трилобитообразные
Класс: Trilobita
Вальч,
заказы
Agnostida
Asaphida
Corynexochida
Harpetida
Nectaspida
Redlichiida
Lichida
Odontopleurida
Phacopida
Proetida
Ptychopariida
Трилобиты, что означает «три доли», представляют собой ископаемую группу вымерших морских арахноморфных членистоногих, которые образуют класс Trilobita. Трилобиты образуют одну из ранних известных групп членистоногих. Первое появление трилобитов в летописи окаменелостей определяет основу атдабановской стадии раннего кембрийского периода (521 млн. Лет назад), и они процветали в течение более низкой палеозойской эпохи, прежде чем начать затянувшийся упадок до исчезновения, когда во время девона, Все трилобитные заказы, кроме проэфидов, вымерли. Трилобиты исчезли в массовом вымирании в конце перми около 252 миллионов лет назад. Трилобиты были одними из самых успешных из всех ранних животных, бродивших океаны более 270 миллионов лет .
К тому времени, когда трилобиты впервые появились в летописи окаменелостей, они были уже сильно диверсифицированы и географически разбросаны. Поскольку трилобиты имели широкое разнообразие и легко окаменевший экзоскелет, обширная летопись окаменелостей осталась позади, а около 17 000 известных видов охватывали палеозойское время. Изучение этих окаменелостей способствовало существенному вкладу в биостратиграфию, палеонтологию, эволюционную биологию и тектонику плит. Трилобиты часто помещаются в членистоногие субфилы Шизорамия в суперклассе Arachnomorpha (что эквивалентно Arachnata), хотя в литературе найдено несколько альтернативных таксономий.
У трилобитов было много образа жизни; Некоторые двигались над морским дном, как хищники, мусорщики или фильтраторы, и некоторые плавали, питаясь планктоном. Большинство видов жизни, ожидаемых от современных морских членистоногих, встречаются в трилобитах, за исключением, возможно, паразитизма (научные дискуссии продолжаются). Некоторые трилобиты (в частности, семейные олениды), как считается, также развили симбиотические отношения с серосодержащими бактериями, из которых они получали пищу.
1 Ископаемая запись
1.1 Происхождение
1.2 Дивергенция и исчезновение
1.2.1 Эволюционные тенденции
1.2.2 Докембрий
1.2.3 кембрия
1.2.4 ордовика
1.2.5 Силурские и девонские
1.2.6. Каменноугольные и пермские
1.2.7 Окончательное исчезновение
2 Распределение ископаемых
3 Важность
4 Систематика
4.1. Связь с другими таксонами
5 Морфология
5.1 Терминология
5,2 Cephalon
5.2.1 Лицевые швы
5.2.1.1 Спинные швы
5.2.1.2 Вентральные швы
5.2.2 Трибуна
5.2.3 гипостомы
5,3 Туловище
5,4 Пигидий
5.5 Просопон (поверхностная скульптура)
5.6 Шипы
6 Мягкие части тела
6.1 придатков
6.2. Пищеварительный тракт
6.3 Внутренние органы
6,4 Мышцы
7 Сенсорные органы
7,1 Усики
7.2 Глаза
7.3 Сенсорные ямы
8 Разработка
Redlichiida, такой как этот Paradoxides, может представлять родовые трилобиты.
Walliserops trifurcatus, от Джебель Оуфатен, Марокко
Самые ранние трилобиты, известные из летописи окаменелостей, - это краснолицы и питопаридные биготиниды, датированные примерно от 540 до 520 миллионов лет назад. Претендентами на самые ранние трилобиты являются Profallotaspis jakutensis (Сибирь), Fritzaspis spp. (Западная часть США), Hupetina antiqua (Марокко) и Serrania gordaensis (Испания). Предполагается, что все трилобиты возникли в современной Сибири с последующим распределением и излучением из этого места.
У всех Оленеллины нет лицевых швов, и это считается оригинальным состоянием. Самый ранний сшитый трилобит, найденный до сих пор (Lemdadella), встречается почти одновременно с самой ранней Оленеллиной, однако, подразумевая, что начало трилобита лежит перед началом атдабани, но не оставляет окаменелостей. Другие группы показывают вторичные потерянные лицевые швы, такие как все Agnostina и некоторые Phacopina. Еще одна общая особенность Оленеллины также предполагает, что этот подотряс является запасом трилобита предков: ранние стадии протопипида не были найдены, предположительно потому, что они не были кальцифицированы, и это также должно представлять собой исходное состояние . Более ранние трилобиты могут быть найдены и могут пролить больше света на происхождение трилобитов.
Три экземпляра трилобита из Марокко Megistaspis hammondi, датированные 478 миллионами лет, содержат окаменелые мягкие части.
Происхождение
Ранние трилобиты показывают все особенности трилобитовой группы в целом; Переходные или предковые формы, показывающие или сочетающие особенности трилобитов с другими группами (например, ранние членистоногие), как представляется, не существую Сходство между трилобитами и ранними членистоногими существами, такими как Сприггина, Парванкорина и другие «трилобитоморфы» периода Эдиакарана докембрия, достаточно неоднозначны, чтобы сделать детальный анализ их предков далеко не убедительными. Морфологическое сходство между ранними трилобитами и другими кембрийскими членистоногими (например, фауна Берджесского сланца и фауна майояншанских сланцев) затрудняет анализ отношений предков. Однако, что трилобиты разделяют общего предка с другими членистоногими, прежде чем граница Эдиакаран-кембрия все еще разумна. Доказательства свидетельствуют о том, что значительная диверсификация уже произошла до того, как трилобиты были сохранены в летописи окаменелостей, что легко позволяет «внезапное» появление различных групп трилобитов со сложными производными характеристиками (например, глазами). Дивергенция и вымирание [редактировать] Olenoides serratus. Кровати Стивена Трилобита (средний кембрий) возле поля, Британская Колумбия, Канада. Для такой многолетней группы животных неудивительно, что история эволюции трилобитов отмечена рядом событий исчезновения, когда некоторые группы погибли, в то время как выжившие группы были диверсифицированы. Заполняют экологические ниши сопоставимыми или новыми приспособлениями. Как правило, трилобиты сохраняли высокий уровень разброса на протяжении кембрийского и ордовикского периодов, прежде чем вступать в затянувшийся спад в девоне, кульминацией которого стало окончательное исчезновение последних выживших в конце пермского периода. Эволюционные тенденции. Основные эволюционные Тенденции из примитивных морфологий, такие как, например, Eoredlichia, включают в себя происхождение новых типов глаз, улучшение механизмов регистрации и артикуляции, увеличение размера пигидия (микропигмия до изопиггии) и развитие крайней спинозиции в определенных группах. Изменения также включали сужение грудной клетки и увеличение или уменьшение количества грудных сегментов. Также отмечаются специфические изменения в цефалоне; Переменный размер и форму глабели, положение глаз и лицевых швов и специализация гипостома. Несколько морфологий проявлялись независимо в разных основных таксонах (например, для уменьшения или миниатюризации глаз). Обесцвечивание, потеря поверхностных деталей в цефалоне, пигидии или грудных бороздах также является общей эволюционной тенденцией. Примечательными примерами этого были приказания Агностида и Асафида и подотряда Илланина Коринексохиды. Считается, что отвращение является показателем либо рогового образа жизни, либо пелагического. Угроза представляет проблему для таксономистов, поскольку потеря деталей (особенно глабели) может затруднить определение филогенетических отношений. Докембрийский филогенетический биогеографический анализ раннего кембрия Olenellidae и Redlichiidae предполагает, что однородная фауна трилобитов Существовали над Лаврентия, Гондвана и Сибири до тектонического разлома суперконтинента Паннотия между 600 и 550 миллионами лет назад. Затем тектонический распад Паннотии позволил диверсифицировать и излучение, выраженное позднее в кембрии, как отличительная провинция оленеллидов (Лаврентия, Сибирь и Балтика) и отдельная провинция краснолитов (Австралия, Антарктида и Китай). Распад Pannotia значительно предшествует первому появлению трилобитов в летописи окаменелостей, поддерживая длительное и загадочное развитие трилобитов, простирающихся, возможно, еще на 700 миллионов лет назад или, возможно, еще вовремя назад. Cambrian Очень скоро Трилобитовые окаменелости появились в нижнем кембрии, они быстро диверсифицировались в основные приказы, типичные для кембрийско-редлихейдских, птихопаридских, агностидских и коринексохид. Первый крупный кризис в летописи ископаемых трилобитов произошел в Среднем Кембрии; Сохранившиеся заказы развивали тела изопигуса или макропигиуса и развивали более толстые кутикулы, позволяя лучше защищать от хищников. Конечное кембрийское массовое вымирание ознаменовало значительное изменение фауны трилобитов; Почти все Redlichiida (включая Olenelloidea) и большинство запасов позднего кембрия вымерли. Продолжающееся снижение площади лавонтовского континентального шельфа регистрируется одновременно с вымираниями, что указывает на серьезные экологические потрясения. В родах, которые появляются в кембрии, имеются триробитовые роды: Абудиалла (нижний кембрий) Юдомия (нижний кембрий) Оленелл ( Нижний кембрий) Эллипсоцефал (средний кембрий) Эльрафия (средний кембрий) Парадоксиды (средний кембрий) Перонопсис (средний кембрий) Xiuqiella (средний кембрий) Йилангелла (средний кембрий) Йиланджеллина (средний кембрий) Олен (поздний кембрий) Ордовик. В ролях Isotelus Рекс, самый крупный известный трилобит от среднего до верхнего ордовика Северной Америки. Черирус, средний ордовик, река Волхов, Россия. Ранний ордовик характеризуется энергичными излучением суставных брахиопод, мшанок, двустворчатых моллюсков, иглокожих и граней Ptolites, со многими группами, впервые появившимися в летописи окаменелостей. Несмотря на то, что во время кембрия, по-видимому, развилось разновидность трилобитов внутри видов, трилобиты все еще были активными участниками ордовикского радиационного события с новой фауной, захваченной со старой кембрийской. Факопида и тринуклеоида являются характерными формами, сильно дифференцированными и разнообразными, большинство из которых с неопределенными предками. Факопида и другие «новые» клады почти наверняка были кембрийскими предками, но тот факт, что они избегали обнаружения, является убедительным свидетельством того, что новые морфологии развивались очень быстро. Изменения в фауне трилобитов во время ордовика предвещали массовое вымирание в конце ордовика, позволяя многим семьям беспрепятственно продолжать в силуре. Ордовикские трилобиты были успешными при эксплуатации новых условий, в частности рифов. Тем не менее, конец ордовика массового вымирания не оставил трилобиты невредимыми; Некоторые отличительные и ранее успешные формы, такие как Trinucleioidea и Agnostida, вымерли. Ордовик отмечает последний великий период диверсификации среди трилобитов: очень мало совершенно новых моделей организации возникло после ордовик. Более поздняя эволюция в трилобитах была в значительной степени вопросом вариаций ордовиков. При ордовищевом массовом вымирании энергичное излучение трилобита прекратилось, и постепенно появляется угасание постепенного упадка. Некоторые из родов трилобитов, появляющихся в ордовикке, включают: Циклопиг (от раннего до позднего ордовика) Селенопельт (от раннего до позднего ордовика) Параболина (ранний Ордовик) Шерирус (средний ордовик) Эодальманитина (средний ордовик) Тринуклеус (средний ордовик) Триартр (поздний ордовик) Силур и девон. Число семейств, средний девон до верхнепермской. Proetida - коричневый, Phacopida - стальной крем, Lichida - clearblue, Harpetida, Odontopleurida и Corynexochida - розовый, оливковый и фиолетовый. Большинство ранних силурийских семей составляют подгруппу фауны позднего ордовика. Немногие, если они есть, доминирующей раннеордовикской фауны дожили до конца ордовика, но 74% доминирующей позднеордовикской трилобитовой фауны пережили ордовика. Поздноордовикские выжившие составляют все послеордовикские трилобитовые группы, за исключением Harpetida. Силурийские и девонские трилобитовые комплексы внешне сходны с ордовикскими сообществами, в которых доминируют Личида и Факопида (включая известную Калименину). Однако ряд характерных форм далеко не распространяется на девона, и почти все остальные были уничтожены серией резких вымираний среднего и позднего девона. Три порядка и все, кроме пяти семей, были уничтожены сочетанием изменений уровня моря и разрыва в окислительно-восстановительном равновесии (воздействие метеорита также было предложено как причина). Только один порядок, Проетида, выжил в карбоне. Роды трилобитов во время силурийского и девонского периодов включают: Дальманиты (от раннего до позднего силура) Калимен (силурий) Энкринурус (силурий) Эксальпасс (от середины до поздней Силуриан) Параледжуры (ранний девон) Лиоарпес (средний девон) Факопы (от среднего до позднего девона) Каменноугольные и пермские. Проетида выжила в течение миллионов лет, продолжалась через период карбона и продолжалась до конца перми (когда Подавляющее большинство видов на Земле были уничтожены). Неизвестно, почему только заказ Протида пережил девон. Проэтида поддерживала относительно разнообразные фауны в глубокой воде и мелкой воде, шельфовые среды по всему карбону . В течение многих миллионов лет Проетида существовала без проблем в своей экологической нише. Аналогами будут сегодняшние криноиды, которые в основном существуют как виды глубоких вод; В эпоху палеозоя обширные «леса» криноидов жили в мелководных прибрежных средах. Некоторые из родов трилобитов в каменноугольном и пермском периодах включают в себя: Гесслериды архегонуса (от раннего до среднего карбона) (среднекаменноугольные ) Эндопс (среднепермский) Трипоэтус (позднекаменноугольный до ранней перми) Дитомопиг (позднекаменноугольный до поздней перми) Окончательное вымирание. Точно, почему трилобиты вымерли, неясно; С повторными событиями экстинкции (часто с последующим очевидным восстановлением) во всей записи о трифобитных окаменелостях, вероятна комбинация причин. После события исчезновения в конце девонского периода то, что оставалось трилобитом, оставалось узким местом в порядке Proetida. Уменьшение разнообразия родов, ограниченных местообитаниями на мелководье, в сочетании с резким понижением уровня моря (регрессия) означало, что окончательное снижение трилобитов произошло незадолго до окончания пермского массового вымирания. С таким количеством морских видов, участвующих в пермском вымирании, конец почти 300 миллионов успешных лет для трилобита в то время не был бы неожиданным. Ближайший известный e Ксантовыми родственниками трилобитов являются подковообразные крабы. Распределение ископаемых. Круциана, трассировка ископаемого трилобитного борозда Трилобитовый фрагмент (Т) в тонком разрезе ордовикского известняка; Е = иглокожих; Шкала шкалы 2 ммRusophycus, «остаточный след» трилобита; Ордовик южного Огайо. Шкала шкалы составляет 10 мм. Пластинка из работы Барранде. Систюм силурен в центре де ла Бохем. Трилобиты, по-видимому, были исключительно морскими организмами, так как окаменелые остатки трилобитов всегда встречаются в породах, содержащих ископаемые другие животные соленой воды, такие как брахиопод, криноиды, И кораллы. В морской морской среде трилобиты были найдены в широком диапазоне от чрезвычайно мелкой воды до очень глубокой воды. Трилобиты, такие как брахиоподы, криноиды и кораллы, встречаются на всех современных континентах и занимают каждый древний океан, из которого были собраны палеозойские окаменелости. Остатки трилобитов могут варьироваться от сохранившегося тела до кусочков экзоскелета, который он проливает в процессе, известном как шелушение. Кроме того, следы, оставленные трилобитами, обитающими на морском дне, часто сохраняются как следовые ископаемые. Существуют три основные формы следовых окаменелостей, связанных с трилобитами: Русофикус; Cruziana & Diplichnites - такие следовые ископаемые представляют собой сохраненную жизнедеятельность трилобитов, активных на морском дне. Русофикус, следы покоя, - это раскопки трилобитов, в которых мало или вообще нет движения вперед, а этилогические интерпретации предполагают отдых, защиту и охоту. Круциана, следы кормления, представляет собой борозды через осадок, которые, как полагают, представляют собой движение трилобитов при подаче осадка. Считается, что многие из окаменелостей Димитрихнитов являются следами, сделанными трилобитами, идущими по поверхности осадка. Тем не менее, следует проявлять осторожность, поскольку аналогичные следовые ископаемые регистрируются в пресноводных и после палеозойских отложениях, представляющих собой не трилобитовые источники. Окаменелости тиробита встречаются во всем мире со многими тысячами известных видов. Поскольку они появились быстро в геологическое время и линьи, как и другие членистоногие, трилобиты служат прекрасными ископаемыми окаменелостями, что позволяет геологам определить возраст пород, в которых они найдены. Они были одними из первых ископаемых, которые привлекали широкое внимание, и каждый год открываются новые виды. Известное месторасположение ископаемых трилобитов в Соединенном Королевстве - это гнездо Рена, Дадли в Уэст-Мидлендсе, где Calymene blumenbachi находится в Силурианской группе Венлока , Этот трилобит представлен на гербе города и был назван Dudley Bug или Dudley Locust карьерами, которые когда-то работали теперь заброшенными известняковыми карьерами. Лландриндод-Уэллс, Повис, Уэльс, является еще одним известным местом трилобита. Известный Elrathia kingi trilobite встречается в изобилии в кембрийском возрасте Уилер-Шейл штата Юта. В Британской Колумбии были обнаружены эффектно сохранившиеся трилобитовые окаменелости, часто показывающие мягкие части тела (ноги, жабры, усики и т. Д.), Канада (кембрийский бургезский сланцы и аналогичные населенные пункты); Нью-Йорк, США (карьера Ордовика Уолкотта-Руста, недалеко от России, и кровать Трилобита Бичера, недалеко от Рима); Китай (нижний кембрийский маотианшанский сланцы около Чэнцзяна); Германия (девонские хунсрюкские сланцы вблизи Бунденбаха) и, что гораздо реже, в трилобитосодержащих пластах в штате Юта (Уилер Шейл и другие образования), Онтарио и Реке Мануэль, Ньюфаундленде и Лабрадоре. Французский палеонтолог Иоахим Барранде (1799-1883) Выполнил свое историческое исследование трилобитов в кембрии, ордовике и силуре из Богемии, опубликовав первый том Силурье дю Сен-де-ла-Бохема в 1852 году. Изучение палеозойских трилобитов в границах валлийского англичан Нилсом Элдриджем Было основополагающим в формулировании и тестировании прерывистого равновесия как механизма эволюции. Идентификация «Атлантического» и «Тихоокеанского» трилобитовых фаун в Северной Америке и Европе подразумевала закрытие Озера Япета (Создание шва Iapetus), тем самым предоставляя важные подтверждающие доказательства теории дрейфа континента. Трилобиты были важны для оценки скорости видообразования в течение известного периода Как Камбрийское Взрыв, потому что они являются самой разнообразной группой метазоанов, известных из летописи окаменелостей раннего кембрия. Трилобиты - превосходные стратиграфические маркеры кембрийского периода: исследователи, которые находят трилобиты с пищевым просопоном и микропигием , Нашли ранние кембрийские слои. Большая часть кембрийской стратиграфии основана на использовании ископаемых трилобитных маркеров. Трилобиты являются государственными окаменелостями Огайо (Isotelus), Висконсином (Calymene celebra) и Пенсильвания (Phacops rana). См. Также: Список родов трилобитов. Таксономический ранг многих группировок трилобитов может быть получен из суффикса к его названию. В следующей таблице перечислены разные ранги (от самых близких к ближайшим родственникам), с соответствующими Суффикс суффикса и пример. Суффикс к названиям таксономических трилобитовых группировок различного рангарк-суффикса exampleclass -a Trilobitaorder -ida Ptychopariidasuborder -ina Eodiscinasuperfamily -oidea Acastoideafamily -idae Cyclopygidaesubfamily -inae Cybelinae 9 заказов трилобитов: Agnostida, Redlichiida, Corynexochida, Lichida, Phacopida , Proetida, Asaphida, Harpetida и Ptychopariida. Иногда Nektaspida включены, но у них нет кальцинированного экзоскелета и глаз. Другие ученые не считают Агностину трилобитами и, следовательно, не связаны с Eodiscina. Этот последний подотряд должен быть повышен и быть назван Eodiscida. Несмотря на их богатую историю окаменелостей с тысячами описанных родов, обнаруженных во всем мире, таксономия и филогения трилобитов имеют много неопределенностей. За исключением, возможно, для членов приказов Phacopida и Lichida (которые впервые появляются во время раннего ордовика), девять из одиннадцати трилобитных орденов появляются до конца кембрия. Большинство ученых полагают, что порядок Редлихиды, а именно его подотряда Редлихина, содержит общего предка всех других порядков, за исключением, возможно, Агностины. Хотя в литературе обнаружено множество потенциальных филогении, у большинства из них есть подотряда Редлихиина, приводящая к порядкам Corynexochida и Ptychopariida во время нижнего кембрия, а Личида, спускающаяся либо из Redlichiida, либо из Corynexochida в среднем кембрии. Приказ Ptychopariida является наиболее проблематичным порядком классификации трилобитов. В 1959 году «Трактат о беспозвоночной палеонтологии», которые теперь являются членами орденов Птихопариды, Асафиды, Проетиды и Гарпетиды, были сгруппированы в качестве ордена Птихопариды; Подкласс Librostoma был установлен в 1990 году, чтобы охватить все эти приказы, основываясь на их общем родовом характере натанта (непривязанного) гипостома. Самый последний признанный из девяти трилобитских орденов, Harpetida, был установлен в 2002 году. Прародитель порядка Phacopida неясен. Отношения с другими таксонами [править] После того, как анатомия мягкой части была восстановлена, трилобиты первоначально были связаны с ракообразным; Однако это предложение с тех пор не в пользу. Отношения с Chelicerata в кладе, названной Arachnomorpha (Arachnata), были в моде в течение некоторого времени; Положение в группе Мандибулата (= Myriapoda + Crustacea + Hexapoda) может быть более скупой альтернативой. Морфология. Когда найдены трилобиты, сохраняется только экзоскелет (часто в неполном состоянии) во всех, кроме Несколько мест. Несколько мест (Lagerstätten) сохраняют идентифицируемые части мягкого тела (ноги, жабры, мускулатуру и пищеварительный тракт) и загадочные следы других структур (например, мелкие детали структуры глаз), а также экзоскелет. Тилобиты имеют длину от минуты (меньше 3 миллиметра (0,12 дюйма)) до очень больших (более 30 см (12 дюймов)) со средним размером 3-10 см (1,2-3,9 дюйма). Предположительно, самый маленький вид - Acanthopleurella specialulae с максимумом 1,5 миллиметра (0,059 дюйма). В 1998 году канадскими учеными в ордовикских породах на берегах Гудзонова залива был обнаружен самый крупный в мире известный образец трилобита, присвоенный Isotelus rex 72 см. Тело трилобита разделено на три основных участка (тагмата): 1 - Cephalon; 2 - грудная клетка; 3 - пигидий. Трилобиты названы так для трех продольных лепестков: 4 - правая плевральная доля; 5 - осевая доля; 6 - левая плевральная доля; На этих диаграммах не показаны антенны и ноги. Экзоскелет состоит из минералов кальцита и кальция фосфата в решетке хитина, которая покрывает верхнюю поверхность (дорзальный) трилобита и скручивается вокруг нижнего края, чтобы создать небольшую полосу, называемую «внутренняя сторона переплета». Существуют три отличительные тагматы (разделы): цефалон (голова); Грудной клетки (тела) и pygidium (хвост). Терминология [править] Как и следовало ожидать для группы животных, включающей c. 5000 родов, морфология и описание трилобитов могут быть сложными. Однако, несмотря на морфологическую сложность и нечеткое положение в высших классификациях, существует ряд характеристик, которые отличают трилобиты от других членистоногих: обычно субэллиптический, дорсальный, хитинозный экзоскелет, разделенный продольно на три отдельные лепестки (из которых группа получает свои имя); Имеющий четкий относительно большой головной щиток (цефалон), шарнирно сочлененный в осевом направлении грудной клеткой, состоящей из сочлененных поперечных сегментов, задняя часть которых почти всегда слита с образованием хвостового щитка (pygidium). При описании различий между таксонами трилобита часто упоминаются наличие, размер и форма головных функций. Во время линьки экзоскелет обычно разделяется между головой и грудной клеткой, и поэтому так много ископаемых трилобитов отсутствует. В большинстве групп лицевые швы на цефалоне помогли облегчить линьку. Подобно омарам и крабам, трилобиты будут иметь физику «вырос» между стадией линьки и упрочнением нового экзоскелета. Цефалон [править] См. Также: Cephalon (анатомия членистоногих) Морфология трилобитного цефалонаЛаблированная диаграмма основных частей цефалона Основные подразделения цефалона. Подразделения могут быть Далее разбиты на разные области, используемые при описании морфологии трилобитов головного мозга. 1 - предпоследняя зона; 2 - пальпебральная область; 3 - постокулярная область; 4 - заднепроходная проекция; 5 - затылочное кольцо; 6 - глабелла; 7 - задняя область; 8 - боковая граница; 9 - либригенная область; 10 - предглабельная область. Цефалон трилобита или головная часть сильно варьируются с большой морфологической сложностью. Глабелла образует купол, под которым сидел «урожай» или «живот». Обычно экзоскелет имеет мало отличительных вентральных особенностей, но цефалон часто сохраняет шрамы для прикрепления мышц, а иногда и гипостому, небольшую жесткую пластинку, сравнимую с вентральной пластинкой у других членистоногих. Беззубый рот и желудок сидели на гипостоме, рот обращен назад на задний край гипостомы. Гипосомная морфология сильно варьируется; Иногда поддерживаемый неминерализованной мембраной (натантом), иногда слитой в переднюю двойку с контуром, очень похожим на глабелицу выше (conterminant) или слитым с передним удвоением с контуром, значительно отличающимся от глабели (подталкивающим). Было описано много изменений в форме и размещении гипостомы [50]. Размер глабели и боковой челюсти цефалона вместе с вариацией гипостомы были связаны с различными формами жизни, диетами и конкретными экологическими нишами. Передняя и боковая полоса цефалона значительно увеличены в Harpetida, в Другие виды выбухают в предглабельной области, что говорит о мешочке-выводке. Очень сложные составные глаза - еще одна очевидная особенность цефалона. Фальцевальные швы. [Править] Лицевые или головные швы являются естественными линиями перелома в цефалоне трилобитов. Их функция заключалась в том, чтобы помочь трилобиту в проливе его старого экзоскелета во время шелушения (или линьки). Все виды, отнесенные к подотряду Оленеллина, которые вымерли в самом конце раннего кембрия (например, Fallotaspis, Nevadia, Judomia и Olenellus) не хватало лицевых швов. Считается, что они никогда не развивали лицевые швы, предварительно подготовив их эволюцию. Из-за этого (наряду с другими примитивными характеристиками) они считаются самыми ранними предками поздних трилобитов. Некоторые другие поздние трилобиты также потеряли лицевые швы вторично. Тип швов, найденных у разных видов, широко используется в таксономии и филогении трилобитов. Спинные швы. Спинной поверхности трилобитового цефалона (самая ранняя тагма или «голова») можно разделить на две Регионы - кранидий и либригена («свободные щеки»). Кранидий может быть далее разделен на глабели (центральная доля в цефалоне) и фиксигена («фиксированные щеки»). Лицевые швы лежат вдоль передней кромки, при разделении между кранидием и либригеном. Тиробитовые лицевые швы на дорзальной стороне можно условно разделить на пять основных типов, в зависимости от того, где швы заканчиваются относительно генального угла (края, где Боковые и задние края цефалона сходятся). Отсутствует - Лицевые швы отсутствуют в Оленеллине. Это считается примитивным состоянием и всегда сочетается с наличием глаз. Пропариант - лицевой шов заканчивается перед половым углом вдоль бокового края. Примеры родов, показывающих этот тип шва, включают в себя далманиты факопина (Phacopida) и Ekwipagetia of Eodiscina (Agnostida). Гонопатар - лицевой шов заканчивается на кончике генного угла. Примеры родов, демонстрирующих этот тип шва, включают Calymene и Trimerus of Calymenina (Phacopida). Opisthoparian - лицевой шов заканчивается у заднего края цефалона. Примеры родов, показывающих этот тип шва, включают Пелтура Оленина (Птихопарида) и Бумаст Иллаенина (Corynexochida). Это самый распространенный тип лицевого шва. Гипопарический или маргинальный. В некоторых трилобитах спинные швы могут быть вторично потеряны. Несколько образцовых временных рядов видов показывают «миграцию» дорзального шва до тех пор, пока он не совпадает с краями цефалона. Поскольку зрительная поверхность глаза находится на уменьшающейся свободной щеке (или либригене), количество линз имеет тенденцию опускаться, и в конечном итоге глаз исчезает. Потеря спинных швов может возникать из пропарианского состояния, например, в некоторых эодицинах, таких как Веймутия, все Агностина и некоторые Факопина, такие как Дуктина. Однако маргинальные швы, проявляемые арпептидами и тринуклеоидами, получены из опистопаровых швов. С другой стороны, слепота не всегда сопровождается потерей лицевых швов. Тиробитовые лицевые шовные типы. Png Первобытное состояние спинных швов Пропара. Штаммы Opisthoparian развивались несколько раз независимо. Нет примеров пропаральных швов, развивающихся в таксонах с опистопарианской родословной. Трилобиты, которые демонстрируют опистопарные швы, как взрослые, обычно имеют пропаральные швы как возрасты (известные исключения - Юньнаноцефал и Дюйнаспис). Гипопарные швы также возникали независимо в нескольких группах трилобитов. Курс лицевых швов с лицевой стороны зрительной поверхности изменяется, по меньшей мере, настолько же силен, как и в задней ветви, но отсутствие четкой контрольной точки, сходной с геномной Угол затрудняет категоризацию. Одно из наиболее выраженных состояний заключается в том, что передняя часть лицевой швы не обрезает латеральную или лобную границу сама по себе, а совпадает перед глабели и вырезает фронтальную границу на средней линии. Это, в частности, случай в Асафиде. Еще более выраженная ситуация заключается в том, что лобные ветви лицевых швов заканчиваются друг с другом, что приводит к свободным щекам. Это известно у триартруса и у Phacopidae, но в этом семействе лицевые швы не являются функциональными, что можно сделать из того факта, что свободные щеки не найдены отделенными от кранидия. Существуют также два типа швов на спинном поверхность, соединенная с составными глазами трилобитов.
Это: Глазные швы - это швы, окружающие края сложного глаза. Трилобиты с этими швами теряют всю поверхность глаз при линьке. Это обычное явление среди кэмбрийских трилобитов. Пальпебральные швы - это швы, которые составляют часть спинного лицевого шва, проходящего по верхним краям составного глаза. Вентральные швы. Трилобитовый цефалон вентральной анатомии.png Дорсальные лицевые швы продолжают вниз к вентральной стороне Цефалон, где они становятся связующими швами, которые делят двойственность. Ниже перечислены типы вентральных швов. Соединительные швы - это швы, которые продолжаются от лицевых швов за передним краем цефалона. Ровный шов - присутствует только тогда, когда трилобит имеет трибуну (или ростральную пластинку). Он соединяет трибуну с передней частью дорсального кранидия. Гипостомальный шов - отделяет гипостому от двойственности, когда гипостома относится к прикрепленному типу. Он отсутствует, когда гипостома является свободно плавающей (т.е. натальной). Он также отсутствует в некоторых котерминантных гипостомах, где гипостома слита с удвоением. Шов меда - проявляемый асафидными трилобитами, они образуются, когда вместо того, чтобы стать соединительными швами, два спинных шва сходятся в точке перед цефалоном, затем делят Прямо вниз по центру удвоения. Рострум (или ростральная пластинка) представляет собой отдельную часть двойственности, расположенную в передней части цефалона. Он отделен от остальной части удвоения ростральным швом. Во время линьки в трилобитах, таких как парадоксиды, рострум используется для закрепления передней части трилобита, когда кранидий отделяется от либригена. Отверстие, созданное выгибанием тела, обеспечивает выход для линьки трилобита. В некоторых трилобитах, таких как Lachnostoma.Hypostome, отсутствует выход. Дополнительная информация: гипостома (трилобит). Асафус экспансус вентральной стороны подготовлен, показывая прикрепление гипостомы. Гипостома Представляет собой твердую часть трилобита, обнаруженную на вентральной стороне цефалона, как правило, ниже глабели. Гипостому можно разделить на три типа, основываясь на том, постоянно ли они прикреплены к трибуне или нет и совпадают ли они с передним дорзальным кончиком глабели. Натант - гипотома, не привязанная к удвоению. Совместим с передним краем glabella.Conterminant - Hypostome прикрепляется к ростральной пластине двойного действия. Совместим с передним краем glabella.Impendent - Hypostome прикреплен к ростральной пластинке, но не выровнен к glabella. Below - иллюстрация трех типов. Двукратное проявление светло-серого цвета, внутренняя поверхность цефалона темно-серого цвета и гипостома светло-голубая. Глабелла очерчена красными ломаными линиями. Типы гипостомов тиробитов на основе прикрепления. Thorax Зачисленный phacopid trilobite Flexiclaymene meeki от верхнего ордовика Огайо. Грудная клетка представляет собой серию сочлененных сегментов, которые лежат между цефалоном и пигидий. Количество сегментов колеблется между 2 и 103 с большинством видов в диапазоне от 2 до 16. Каждый сегмент состоит из центрального осевого кольца и наружных плевр, которые защищают конечности и жабры. Плеверии иногда сокращаются или расширяются, образуя длинные шипы. Apodemes - это луковичные выступы на вентральной поверхности экзоскелета, к которому прикреплены большинство мышц ног, хотя некоторые мышцы ног прикреплены непосредственно к экзоскелетону. Определение соединения грудной клетки и пигидия может быть затруднено, и многие из них страдают от этой проблемы. Ископаемые трилобиты часто встречаются «зачисленными» (свернувшись), как современные таблетки для ащита; Свидетельства свидетельствуют о том, что регистрация помогла защитить от присущей слабости кутикулы членистоногих, которую использовали аномалокаридные хищники. Некоторые трилобиты достигли полностью закрытой капсулы (например, Phacops), тогда как другие с длинными плевральными шипами (например, Selenopeltis) оставили пробел в Стороны или те, у кого небольшой пигидий (например, парадоксиды), оставили разрыв между цефалоном и пигидием. В Phacops плевры перекрывают гладкий скос (грань), позволяя закрыть уплотнение с удвоением. На двойном удлинении на каждом сегменте имеется выемка или выемка Пандериана для предотвращения вращения и достижения хорошего уплотнения. Даже в агностисте, только с 2-мя артикуляционными грудными сегментами, процесс зачисления требовал сложной мускулатуры, чтобы сжимать экзоскелет и возвращаться в плоское состояние. Пигидий образуется из ряда сегментов и тельсона Слитые вместе. Сегменты в pygidium похожи на грудные сегменты (несущие бирамальные конечности), но не сочленены. Трилобиты могут быть описаны на основе микропигуса пигидия (пигидия меньше цефалона), субизопигового (пигидийного субцефана, равного цефалону), изопиготного (пигидий, равного по размеру цефалону), или макропиготного (пигидий больше цефалона) .Прозопон (поверхностная скульптура) Koneprusia brutoni, пример вида с развитыми шипами из девонской хамарской лахдадской свиты, Alnif, MoroccoTrilobite exoskeletons показывают множество мелких структур, которые все вместе называются просопоном. Prosopon не включает крупномасштабные удлинения кутикулы (например, полые плевральные иглы), но для более мелких функций, таких как ребра, купола, пустулы, питтинг, гребень и перфорации. Точная цель просопона не решена, но предложения включают структурное укрепление, сенсорные ямы или волосы, предотвращение нападений хищников и поддержание аэрации во время регистрации. В одном примере пищевые хребтовые сети (легко видны в кэмбрийских трилобитах) могли быть либо пищеварительными, либо респираторными трубками в цефалоне и в других регионах. Спины Некоторые трилобиты, такие как порядок Личиды, развивали сложные колючие формы, От ордовика до конца девонского периода. Примеры этих образцов были обнаружены в Хамарской Лагдадской свите Альнифа в Марокко. Тем не менее, существует серьезная проблема подделки и фальсификации большей части марокканского материала, предлагаемого на коммерческой основе. Захватывающие трилобиты также обнаружены на западе России; Оклахома, США; И Онтарио, Канада. У некоторых трилобитов были рога на голове, похожие на несколько современных жуков. Исходя из размера, местоположения и формы рогов, было высказано предположение, что эти рога могли использоваться для борьбы с товарищами. Рога были широко распространены в семье Raphiophoridae (Асафида). Другой функцией этих шипов была защита от хищников. При зачислении трилобитовые шипы предлагали дополнительную защиту. Этот вывод, вероятно, применим и к другим трилобитам, например, к тиробиту Phacopid Walliserops, который развивал захватывающие трезубцы. Мягкие части тела/ Описаны только 21 вид, из которого сохраняются мягкие части тела , поэтому некоторые особенности (например, задние антеннообразные церки, сохранившиеся только у Оленоидов серратуса) по-прежнему трудно оценить в более широкой картине. реконструкция жизни Triarthrus eatoni на основе сохраненных частей мягкого тела. Рисунок бирамальной ноги Agnostus pisiformisВентральная сторона Triarthrus eatoni, показывающая антенны и бирамальные ноги, сохраненные в пирите. Исключительно хорошо сохранившийся Olenoides serratus из Burgess Shale. Антенны и ноги сохраняются как отражающая углеродная пленка. Тиробиты имеют одну пару предраковых антенн и в остальном недифференцированные бирамальные конечности (2, 3 или 4 цефальные пары, за которыми следуют одна пара на грудной сегмент и некоторые пары пигидия). Каждый эндоподит (ходьба) имел 6 или 7 сегментов, гомологичный другим ранним членистоногим. Эндоподиты прикреплены к коксам, которые также имели периподобный экзоподит или жаберную ветвь, которая использовалась для дыхания и у некоторых видов плавание. Внутренняя часть coxa (или gnathobase) несет шипы, вероятно, для обработки предметов жертвы. Последний сегмент экзоподита обычно имел когти или шипы. Многие примеры волосков на ногах предполагают адаптацию для кормления (как для гнатобаз), так и для сенсорных органов, чтобы помочь в ходьбе. Пищеварительный тракт. Беззубый рот трилобитов находился на заднем крае гипостомы (обращенный назад ), Перед ногами, прикрепленными к цефалону. Рот связан небольшим пищеводом с желудком, который лежит впереди рта, ниже глабели. «Кишечник» отступил назад к пигидию. Считается, что «кормящие конечности», прикрепленные к цефалону, кормили пищей в рот, возможно, сначала «нарезая» еду на гипостоме и / или гнатобазах. Альтернатива. Предполагается, что при использовании яичников цефальные лапы могут потревожить осадок, чтобы сделать пищу доступной. Большая глабели, (подразумевая большой желудок), в сочетании с подкрепленным гипостомом, использовалась в качестве доказательства более сложных источников пищи, то есть, возможно, плотоядного образа жизни. Внутренние органы. Несмотря на наличие прямых и подразумеваемых доказательств для Наличие и расположение полости рта, желудка и пищеварительного тракта, присутствие сердца, мозга и печени подразумевается (хотя и «присутствует» во многих реконструкциях) с небольшими прямыми геологическими данными. Мускулатура. Хотя редко Длинные боковые мускулы, вытянутые от цефалона до середины пигидия, прикрепленные к осевым кольцам, позволяющие регистрироваться, в то время как отдельные мышцы на ногах спрятали их в сторону. Сенсорные органы. Многие трилобиты имели сложные глаза; У них также была пара антенн. Некоторые трилобиты были слепыми, вероятно, они жили слишком глубоко в море, чтобы свет мог их достигать. Таким образом, они стали вторично слепыми в этой ветви эволюции трилобитов. Другие трилобиты (например, Phacops rana и Erbenochile erbeni) имели большие глаза, которые использовались в более хорошо освещенных, наполненных хищниками водах. Антенна Пара антенн, подозреваемых в большинстве трилобитов (и сохранившихся в нескольких примерах), была очень гибкой Чтобы они были убраны, когда был зарегистрирован трилобит. Кроме того, один вид (Olenoides serratus) сохраняет антенноподобные церки, которые выступают из задней части трилобита. Глаза. Даже самые ранние трилобиты имели сложные глаза с линзами из кальцита (характерные для всех трилобитовых глаз ), Подтверждая, что глаза членистоногих и, возможно, других животных могли развиться до кембрия. Улучшение зрения как хищника, так и добычи в морской среде было предложено как одно из эволюционных давлений, способствующих явному быстрому развитию новых форм жизни во время так называемого Камбрийского Взрыва. Глаза Трилобита обычно были сложными, причем каждая линза была Удлиненную призму. Количество линз в таком глазу варьировалось: у некоторых трилобитов было только одно, в то время как у некоторых были тысячи линз в одном глазу. В сложных глазах линзы обычно располагались в гексагональном положении. Обследование окаменелостей глаз трилобитов достаточно полно, что их эволюция может быть исследована во времени, что в некоторой степени компенсирует отсутствие сохранения мягких внутренних частей. Объективы глаз трилобитов были сделаны из кальцита (карбонат кальция, CaCO3) , Чистые формы кальцита прозрачны, а некоторые трилобиты используют кристаллографически ориентированные, прозрачные кристаллы кальцита для образования каждой линзы каждого из их глаз. Жесткие кальцитовые линзы не могли бы соответствовать изменению фокуса, как мягкая линза в человеческом глазу; Однако в некоторых трилобитах кальцит сформировал внутреннюю дублетную структуру, дающую превосходную глубину поля и минимальную сферическую аберрацию, согласно оптическим принципам, обнаруженным французским ученым Рене Декартсом и голландским физиком Кристианом Гюйгенсом в 17 веке. Живым существом со сходными линзами является хрупкая звезда Ophiocoma wendtii. В других трилобитах, где, по-видимому, отсутствует интерфейс Гюйгенса, применяется индексная градиентная линза с коэффициентом преломления линзы, изменяющимся к центру. Сенсорный сенсор Структуры были обнаружены в глазах некоторых факопидных трилобитов. Структуры состоят из нескольких сенсорных клеток, окружающих радомерную структуру, напоминающих сублисарную структуру, обнаруженную в глазах многих современных глаз аппетитных членистоногих, особенно Лимула, рода подковы крабов. Голохроальные глаза имели большое количество (Иногда более 15 000) небольших (30-100 мкм, редко больших) линз. Линзы были шестигранно плотно упакованы, касаясь друг друга, с одной мембраной роговицы, покрывающей все линзы. Каждая линза находилась в непосредственном контакте с соседними линзами. Голохроальные глаза - это предковый глаз трилобитов и, безусловно, наиболее распространены, встречаются во всех порядках, кроме Агностиды, и во всем существовании трилобитов. Мало что известно о ранней истории голохроальных глаз; Трилобиты нижнего и среднего кембрия редко сохраняют визуальную поверхность. Пространственная разрешающая способность тертых глаз (таких как голохроальные глаза) зависит от интенсивности света, кругового движения, плотности рецептора, зарегистрированного угла света и степени, в которой скрещивается сигнал отдельных рабдомов. Это означает, что линзы должны быть больше в условиях низкой освещенности (например, для Pricyclopyge, при сравнении с каролинитами) и для быстро движущихся хищников и жертв. Поскольку круговая скорость, вызванная прямой скоростью самого животного, намного выше для омматидий, направленных перпендикулярно движению, быстро движущиеся трилобиты (такие как каролиниты) имеют глаза, сплющенные сбоку, и более изогнутые были омматиями направлены на фронт Или назад. Таким образом, Морфология может быть использована для предположения об экосистеме трилобитов. Шизохромный глаз Эрбенохиля erbenii; Глазной тень является однозначным доказательством того, что некоторые трилобиты были сутками. Глаза Шизохроала обычно имели меньше (около 700), более крупные линзы, чем голохроальные глаза, и встречаются только в Факопине. У каждой линзы была роговица, а соседние линзы были разделены толстой межзунной кутикулой, известной как склера. Глаза Шизохруаля появляются внезапно у раннего ордовика и предположительно были получены от голохроального предка. Поле зрения (все вокруг зрения), размещение глаз и случайное развитие более эффективных механизмов зачисления указывают на глаз как на более защитную систему раннего предупреждения, чем на непосредственную помощь в охоте за едой. Современные глаза, функционально эквивалентные шизохральному глазу, не считались существующими, но встречаются у современных видов насекомых Xenos peckii. Глаза абатохроалов встречаются только в кембрийской эодиске и имеют около 70 небольших отдельных линз, которые Имели отдельную роговицу. Склера была отделена от роговицы и была не такой толстой, как склера в глазах шизохроалов. Хотя хорошо сохранившиеся примеры редко встречаются в ранней летописи окаменелостей, в нижнем кембрии зафиксированы абатохоральные глаза, что делает их одними из самых старых. Условия окружающей среды, по-видимому, привели к более поздней потере зрительных органов во многих эодицинах. Вторичная слепота не является редкостью, особенно у долгоживущих групп, таких как Agnostida и Trinucleioidea. В Proetida и Phacopina из Западной Европы и особенно Tropidocoryphinae из Франции (где имеется хороший стратиграфический контроль) хорошо изучены тенденции, свидетельствующие о постепенном сокращении глаз между близкородственными видами, что в конечном итоге приводит к слепоте. Несколько других структур на трилобитах были Объясняется как фоторецепторы. Особый интерес представляют «макула», небольшие участки разреженной кутикулы на нижней стороне гипостомы. В некоторых трилобитах макула предлагается функционировать как простые «вентральные глаза», которые могли бы обнаружить ночь и день или позволили трилобиту перемещаться во время плавания (или поворота) вверх тормашками. Пятнистая бахрома на цефалоне Trinucleoidea trilobiteSensory pits. Существует несколько типов просопонов, которые были предложены в качестве сенсорного аппарата для сбора химических или вибрационных сигналов. Связь между большими ямными полосками на цефалоне Harpetida и Trinucleoidea с соответствующими небольшими или отсутствующими глазами делает интересной возможность бахромы как «сложного уха». Развитие Пять этапов в развитии Sao hirsutaTrilobites выросли Через последовательные стадии линьки, называемые стадиями, в которых существующие сегменты увеличивались по размеру, а новые сегменты стволов появились в субтерминальной генеративной зоне в течение анаморфотной фазы развития. За этим последовала эпиморфная фаза развития, в которой животное продолжало расти и линько, но никаких новых сегментов ствола не было выражено в экзоскелетоне. Сочетание анаморфического и эпиморфного роста представляет собой гемианамфорический способ развития, который является общим для многих живых членистоногих. Развитие трилобитов было необычным в том, как развивались суставы между сегментами, а изменения в развитии артикуляции приводили к общепризнанным Фазы развития трилобитного жизненного цикла (разделенные на 3 стадии), которые не сравнимы с таковыми у других членистоногих. Фактический рост и изменение внешней формы трилобита имели бы место, когда трилобит был мягким шелушением, после линьки и до того, как следующий экзоскелет затвердел. [88] Серия роста Elrathia kingii с холаспидами от 16,2 мм до 39,8 мм в длину Известны личинки тиробита От кембрия до карбона и от всех подрядов. Поскольку возрастные группы близких таксонов больше похожи на возрасты от дистанционно связанных таксонов, личинки трилобитов обеспечивают морфологическую информацию, важную для оценки филогенетических отношений высокого уровня среди трилобитов. Несмотря на отсутствие подтверждающих окаменелостей, их сходство с живыми членистоногими привело к К убеждению, что трилобиты размножаются половым путем и продуцируют яйца. Некоторые виды, возможно, содержали яйца или личинки в мешке-выводке перед глабелицей, особенно когда экологическая ниша вызывала личинки. Размер и морфология первой стадии кальцинирования сильно варьируются между (но не внутри) трилобитными таксонами, что указывает на то, что некоторые трилобиты проходят через рост в яйце, чем другие. Ранние стадии развития до кальцинирования экзоскелета - это возможность (предложенная для fallotaspids), но так же как и кальцификация и вылупление совпадают. Ранняя стадия роста после эмбрионального трилобита, известная с уверенностью, является «протоспидными» стадиями ( Анаморфную фазу). Начиная с неразличимого Oto-cephalon и proto-pygidium (anaprotaspid) происходит ряд изменений, заканчивающихся поперечной бороздой, разделяющей протоцефалон и протопигидий (метапротаспид), которые могут продолжать добавлять сегменты. Сегменты добавляются в заднюю часть пигидия, но все сегменты остаются слитыми. «Мераспидальные» стадии (анаморфная фаза) отмечены появлением артикуляции между головой и слитым сундуком. До начала первой стадии меразипида животное имело двухчастную структуру - головку и пластину саржевых сегментов ствола, пигидий. Во время мерцательных этапов появились новые сегменты у задней части пигидия, а также дополнительные суставы, развивающиеся у передней части пигидия, высвобождающие свободно сочленяющиеся сегменты в грудную клетку. Сегменты обычно добавляются по одному на линьку (хотя также регистрируются две на линь и каждая каждая черепаха), причем количество этапов равно числу грудных сегментов. Значительный рост составил от 25% до 30% -40%, вероятно, на разных этапах. «Голаспид» (эпиморфная фаза) начинается, когда стабильное, зрелое число сегментов Выпущенный в грудную клетку. Линька продолжалась во время голографических стадий, без изменений в числе торакальных сегментов. Предполагается, что некоторые трилобиты продолжают линять и расти на протяжении всей жизни человека, хотя и более медленными темпами, достигающими зрелости. Некоторые трилобиты проявляли заметный переход в морфологии на одном конкретном этапе, который был назван «метаморфизацией трилобитов». Радикальное изменение морфологии связано с потерей или природой отличительных признаков, которые отмечают изменение в образе жизни. Изменение образа жизни во время развития имеет значение с точки зрения эволюционного давления, поскольку трилобит может пройти через несколько экологических ниш на пути к развитию взрослых, и изменения будут сильно влиять на выживаемость и рассеивание таксонов трилобита [88]. Стоит отметить, что трилобиты со всеми протопидными стадиями исключительно планктонных и более поздних меласпидных стадий бентических (например, асафидов) не смогли продержаться через ордовикские вымирания, тогда как трилобиты, которые были планктоническими только для первой стадии протосипида, до того, как метаморфозы в донные формы сохранились (например, личиды, phacopids). Пелагический личиночный образ жизни оказался плохо адаптированным к быстрому началу глобального климатического охлаждения и утраты тропических шельфовых местообитаний во время ордовика. История использования и исследований [редактировать] Рисунок Ogygiocarella debuchii Ред. Эдвардом Ллойдом, сделанный в 1698Rev. Эдвард Ллойд опубликовал в 1698 году в «Философских трудах Королевского общества», старейший научный журнал на английском языке, часть его письма «О нескольких недавно заведомо исчисленных камнями», сопровождавшееся страницей офортов окаменелостей. Одна из его гравюр показала трилобит, который он нашел возле Лландейло, вероятно, на основании замка лорда Динфорда, он описал как «... скелет какой-то плоской рыбы ...». Открытие Калимен-блюмбачи (саранчи Дадли ) В 1749 году Чарльзом Литлтоном, можно было бы назвать началом исследования трилобитов. Lyttleton подал письмо в Королевское общество Лондона в 1750 году относительно «окаменевшего насекомого», которое он обнаружил в «ямах известняка в Дадли». В 1754 году Мануэль Мендес да Коста заявил, что саранча Дадли не была насекомым, а вместо этого принадлежала к «раковому племени животных». Он предложил называть экземпляры Дадли педикюла маринуса майора трилобо (крупную трехлопастую морскую вошь), имя, которое Продолжался и в 1800-х годах. Немецкий натуралист Йоханн Валх, который провел первое инклюзивное исследование этой группы, предложил использовать название «трилобит». Он счел уместным вывести имя из уникального трехлопастного характера центральной оси и зоны плевры с каждой стороны. Письменные описания трилобитов датируются, возможно, с третьего века до нашей эры и определенно с четвертого века нашей эры. Испанские геологи Эладио Лианьян и Родольфо Гозало утверждают, что некоторые из ископаемых, описанных в греческих и латинских лапидариях, как камень скорпиона, камень жука и камень муравья, относятся к ископаемым трилобита. Менее двусмысленные ссылки на ископаемые трилобитов можно найти в китайских источниках. Ископаемые из кушанского образования северо-восточного Китая ценились как чернильные камни и декоративные изделия. В Новом Свете американские охотники за ископаемыми нашли обильные месторождения Эльрафии-царя в западной части штата Юта в 1860-х годах. До начала 1900-х годов коренные американцы штата Юта носили эти трилобиты, которые они называли пахаве (маленькая водяная ошибка), как амулеты. Отверстие было скучно в голове, а ископаемое было надето на веревке. По словам самих Ute, ожерелья трилобитов защищают от болезней и пуль. В 1931 году Фрэнк Беквит обнаружил доказательства использования тритобитов в Уте. Проезжая через бесплодные земли, он фотографировал два петроглифа, которые, скорее всего, представляют собой трилобиты. На В то же время он осмотрел захоронение неизвестного возраста с пробуренной трилобитовой ископаемой, лежащей в грудной полости мещан. С тех пор трилобитовые амулеты были найдены по всему Большому бассейну, а также в Британской Колумбии и Австралии. В 1880-х годах археологи обнаружили в Гроте-дю-Трилобите («Пещеры Арси-сюр-Каре», Йонна, Франция) Много обработанную окаменелость трилобита, которая была просверлена, как если бы ее носили как кулон. Площадь оккупации, в которой был найден трилобит, датируется пятнадцати тысячами лет. Поскольку кулон был обработан так много, виды трилобита не могут быть определены. Этот тип трилобита не найден вокруг Yonne, поэтому он, возможно, был высоко оценен и продавался из других источников.
Метки для поиска: трилобит, трилобиты фото, трилобиты купить, трилобиты Ленинградской области, купить трилобит, трилобит браслет, трилобиты свидетели эволюции, трилобит цена, глаз трилобита, найти трилобита, трилобит видео, окаменелости трилобитов, трилобит мечехвост, жук трилобит, вымирание трилобитов, трилобиты Ленобласть, трилобиты картинки, вымершие трилобиты, виды трилобитов, трилобиты жили, Волхов трилобиты, Ричард Форти трилобиты свидетели эволюции, когда был расцвет трилобитов, окаменелые трилобиты, отпечатки трилобитов, трилобиты продажа цены, трилобиты палеозой, трилобиты предки, ark survival evolved трилобит, Саблино где трилобиты, ордовикские трилобиты, глаз трилобита которому 380 миллионов лет, трилобиты продажа в СПб, мел и известняк образовались из раковин трилобитов, окаменевший трилобит цена, Марокко купить вывезти трилобитов, Марокко трилобиты, трилобиты Форти.
Трилобит, трилобиты фото, трилобиты купить, трилобиты Ленинградской области, купить трилобит, трилобит цена, трилобиты Ленобласть, трилобиты Путилово, Волхов трилобиты, трилобиты продажа цены, , ордовикские трилобиты, окаменевший трилобит цена, Марокко купить вывезти трилобитов, Марокко трилобиты.






Трилобит на породе. Найден: Марокко.
продано






Трилобит на породе. В идеальной, коллекционной сохранности! Найден: Марокко. Примерно 450 миллионов лет.
продано
Трилобит Asaphus kowalewskii. Очень качественная препарация. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, Путилово, средний ордовик, Волховский горизонт.
продано






Трилобит на породе. Очень качественная препарация! Вид - Asaphus kowalewskii. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, Путилово, средний ордовик, Волховский горизонт.
продано
Крупный трилобит на породе. Очень качественная препарация! Вид - Asaphus lepidurus. Датировка - ордовикский период палеозоя. Ордовик начался примерно 488 млн. лет назад и закончился около 440 млн. лет назад. Найден: Ленинградская область, Путилово, средний ордовик, Волховский горизонт.
продано